Escrito por Anderson Garcia
São os
componentes estruturais e essenciais das proteínas, moléculas biologicamente
ativas de todos os seres vivos. Quimicamente são constituídos por um grupo
amino (N+H3), um grupo carboxila (COOH), um átomo de
hidrogênio (H) e um grupo radical, todos ligados a um carbono α (Figura
1).
Figura 1. Estrutura química comum de todos os aminoácidos. A) Representação
no modelo esfera e bastão. B) Representação na forma estrutural plana.
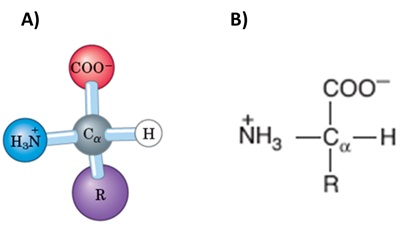
Fonte: NELSON; COX, 2014;
DEVLIN, 2011.
Atualmente,
são conhecidos mais de 300 tipos de aminoácidos encontrados na natureza, mas
destes, somente 20 ocorrem com frequência nas proteínas dos seres vivos, por
isto são tidos e classificados como essenciais. Todos os aminoácidos tidos como essenciais
compartilham dos grupos químicos ilustrados na Figura 1 (exceto o aminoácido
Prolina), porém o que faz com que eles sejam quimicamente e fisicamente distintos
uns dos outros é o seu grupo radical. De acordo com o radical ligado ao carbono
α,
os aminoácidos podem ser classificados como: apolares alifáticos, polares não
carregados, carregados positivamente, carregados negativamente e aromáticos (Figura
2).
Figura 2. Os 20 aminoácidos (essenciais) comumente encontrados nas
proteínas. Classificação de acordo com o grupo R.

Fonte: NELSON; COX,
2014.
É possível notar com clareza na
Figura 2, os grupos químicos comuns à todos os aminoácidos, (exceto a Prolina) e
os grupos R (em róseo) ou também chamados cadeias laterais, que vão diferir os
aminoácidos em suas propriedades físicas e químicas.
Os aminoácidos
pertencentes ao grupo R apolares alifáticos (Glicina, Alanina, Prolina, Valina, Leucina, Isoleucina e Metionina), são hidrofóbicos (insolúveis
em água, pois não são capazes de formar pontes de hidrogênio ou qualquer outro
ter tipo de interação com a mesma). Estes aminoácidos tendem a se posicionar e
alocar seus grupos R no interior das proteínas, longe da água, onde assim,
podem fazer interações hidrofóbicas com outros resíduos de aminoácidos,
contribuindo para a estabilização da estrutura da proteína.
Aqueles que
pertencem ao grupo dos polares, não carregados (Serina, Treonina, Cisteína, Asparagina e Glutamina) são mais hidrofílicos, ou seja, mais solúveis em água
do que aqueles apolares, pois estes contêm grupos funcionais, hidroxila (OH)
(Serina e Treonina), amida (NH2) (Asparagina e Glutamina) e
sulfidrila ou tiol (SH) (Cisteína) que podem fazer ponte de hidrogênio com a
água e contribui para sua polaridade.
Os
pertencentes ao grupo R carregado positivamente, tem cadeias laterais com
grupos amino primário (N+H3) (Lisina), guanidino (Arginina)
e imidazol (Histidina). Estes
grupos R conferem ao aminoácido carga uma carga líquida positiva em pH 7,0,
tornando-os mais hidrofílicos que os outros grupos citados anteriormente.
Os carregados
negativamente são compostos por dois aminoácidos, o Aspartato e o Glutamato,
ambos com um segundo grupo carboxila em sua cadeia lateral
E por fim, o
grupo dos aminoácidos aromáticos, são compostos pelos aminoácidos: Triptofano, Fenilalanina e Tirosina. Todos compartilham de anéis
aromáticos em sua cadeia lateral, e são relativamente apolares. O anel indol do
Triptofano e o grupo hidroxila ligado ao anel aromático da Tirosina faz com que
estes dois aminoácidos sejam pouco apolares quando comparados com a
Fenilalanina que têm somente um anel benzeno em sua cadeia lateral. Devido aos
seus anéis aromáticos, o Triptofano, a Tirosina e em menor extensão a
Fenilalanina, conseguem absorver luz no comprimento de onda de 280 nm (região
UV do espectro).
É importante
conhecer todos os aminoácidos essenciais e seus radicais ou cadeias laterais,
pois estes são responsáveis pela estruturação, hidrofobicidade e interação da
proteína com outras biomoléculas, e também participam como grupo catalítico das
enzimas. Alterações bruscas na sequência de aminoácidos de uma proteína (ex:
substituição de um aminoácido polar por um apolar, ou deleção de uma sequência
específica) podem interferir na sua função, resultando em uma proteína com
propriedades não funcionais, acarretando em alterações na fisiologia e
homeostase do organismo, exemplo: nos casos de anemia falciforme, uma
substituição do aminoácido hidrofílico glutamato por um resíduo hidrofóbico
valina na posição 6 da cadeia β da hemoglobina, faz com que esta
proteína se torne um precipitado fibroso e rígido deformando as hemácias em sua
característica falciforme (forma de foice).
Além dos
aminoácidos essenciais, alguns aminoácidos incomuns estão presentes em algumas
proteínas dos seres vivos, como por exemplo a 4-hidroxiprolina (um aminoácido
derivado da Prolina) e a 5-hidroxilisina (aminoácido derivado da Lisina)
encontradas em proteínas fibrosas como o colágeno, a 6-N-metil-lisina, constituinte da proteína muscular miosina, o γ-carboxiglutamato,
encontrado na proteína trombina e outras que se ligam ao cálcio, a desmosina,
um aminoácido derivado de quatro resíduos de lisina, encontrada na proteína
elastina, a selenocisteína e a cistina, derivada de dois resíduos de Cisteína
unidos por pontes dissulfeto.
Como visto,
alguns aminoácidos não essenciais, podem participar da composição de algumas
proteínas, outros como a Ornitina e Citrulina participam apenas de vias
metabólicas como biossíntese de Arginina e ciclo da uréia.
Propriedades
físico-químicas
Configuração
absoluta
Como
visto anteriormente, os aminoácidos possuem carbono α, onde se ligam os grupos
funcionais que caracterizam esta classe de biomoléculas. O carbono α, exceto
no aminoácido Glicina, é um carbono quiral ou centro quiral, ou seja, possui
quatro ligantes diferentes ligados a ele. Esta propriedade faz com que
os aminoácidos possam ter dois estereoisômeros (moléculas com fórmulas
moleculares iguais, porém com configurações tridimensionais diferentes)
denominados enantiômeros (moléculas com imagens especulares não sobreponíveis
uma a outra).
Para
determinar a configuração absoluta dos aminoácidos e carboidratos, Emil Fisher,
propôs o sistema D e L com base na configuração absoluta do carboidrato de três
carbonos Gliceraldeído (Figura 3.)
Figura 3. Configuração absoluta. Sistema D e L proposto por Emil Fisher,
1981.

Fonte: NELSON; COX,
2014.
Sendo assim, todos aqueles
aminoácidos que tiverem a configuração absoluta relacionada à do
L-Gliceraldeído são tidos como estereoisômeros da série L e aqueles com
configuração relacionada à do D-Gliceraldeído são tidos como estereoisômeros da
série D.
O sistema D e
L diz somente respeito à configuração absoluta e não à atividade óptica da
molécula.
A maioria dos
aminoácidos encontrados na natureza são da série L, com exceção de alguns
D-aminoácidos encontrados em proteínas de paredes celulares de bactérias e
alguns antibióticos.
Em reações
químicas comuns, os aminoácidos podem ser encontrados como uma mistura
racêmica, ou seja, tanto estereoisômeros L quanto D. Nossas células têm a
capacidade de sintetizar somente estereoisômeros da série L.
Ionização
e caráter ácido-básico
Os aminoácidos possuem dois grupos ionizáveis ligados ao
carbono α, o grupo carboxila (COOH) e o grupo amino (N+H3)
e podem ainda, em alguns aminoácidos conter alguns destes grupos na cadeia
lateral (grupos R carregados positivamente e negativamente). Esses grupos podem
ter seus prótons (H+) removidos em função do pH, tornando-se
desprotonados.
A
ionização (perda de prótons) dos seus grupos funcionais e das cadeias laterais,
conferem aos aminoácidos a capacidade de comportarem-se como bases ou ácidos
fracos em função do pH. Podemos dizer então que os aminoácidos são moléculas
anfotéricas, em virtude do seu caráter ácido-básico (Figura 4).
Figura 4. Desprotonação dos grupos funcionais do aminoácido Glicina e suas respectivas cargas em função da perda gradual de prótons.

Fonte: Modificado de
NELSON; COX, 2014.
Figura 5. Formas
não iônicas e zwtteriônicas dos aminoácidos. A forma não iônica
não ocorre em quantidades significativas em soluções aquosas. O zwitteríon
predomina em pH neutro. Um zwitteríon pode atuar tanto como ácido (doador de
prótons) quanto como base (aceptor de prótons) (NELSON; COX, 2014).

Fonte: NELSON; COX, 2014.
Cada aminoácido tem seus grupos funcionais ionizados em
diferentes faixas de pH, o que quer dizer que cada aminoácido tem uma curva de
titulação, valores de pKa
e PI característicos.
* Definições
- ·pKa= é uma medida da tendência de um grupo (COOH e N+H3) doar um próton (H+), com essa tendência diminuindo dez vezes à medida que o pKa aumenta em uma unidade. Um aminoácido diprótico (com dois grupos ionizáveis), podem conter dois pKas, denominados pK1 para a ionização do grupo COOH e pK2 para a ionização do grupo N+H3, e ainda se for um aminoácido triprotico (três grupos ionizáveis), o terceiro grupo é o da cadeia lateral, denominado por pKr. O pKa de um aminoácido é igual ao pH onde ocorre a desprotonação parcial, ou seja, há predominância das duas espécies químicas, a protonada e desprotonada.
- · PI (ponto isoelétrico) = é o ponto onde o aminoácido permanece na forma bipolar ou zwtteriônica.
O
aminoácido glicina, quando titulado com uma base forte, o hidróxido de sódio
(NaOH) produz uma curva característica de onde podem ser extraídas informações
valiosas sobre o comportamento químico deste aminoácido.
Figura 6. Curva
de titulação do aminoácido glicina com uma base forte (NaOH).

Fonte: Modificado de
NELSON; COX, 2014.
De acordo com a Figura 6, é possível observar que em pH
baixo, ente 1,0 e 2,0, a glicina está com seus grupos funcionais totalmente
protonados, conferindo caráter ácido a esta molécula e carga de +1. A medida em
que a titulação segue, a tendência do grupo COOH em doar um H+
aumenta, chegando ao ponto onde, há a ionização deste grupo em um pH de 2,34 (pK1). Neste estágio, a glicina
permanece com metade dos seus grupos carboxila protonados (COOH) e
desprotonados (COO-) com carga relativa de +0,5. No pH de 5,97 ocorre a
desprotonação completa do grupo carboxila, onde a espécie predominante é a N+H3CH2COO-.
Este é o ponto isoelétrico, onde a molécula de glicina permanece como íon
bipolar ou zwetterion, podendo aceitar ou doar prótons e sua carga líquida é 0.
O ponto isoelétrico é calculado a partir da média entre os pKas, ou seja, PI = (pK1+ pK2)/2, no caso de um aminoácido diprótico. Seguindo com a
titulação, o próximo ponto é pK2,
onde há a ionização do grupo amino em pH 9,60, predominando as espécies
protonada (N+H3) e desprotonada (H2N) e sua
carga relativa de -0,5. A titulação está completa em pH 12, onde ocorre a
desprotonação completa do grupo amino, predominando a espécie química NH2CH2COO-
com carga líquida de -1.
A
curva de titulação oferece informações importantes sobre a ionização dos grupos
funcionais e carga da glicina, evidenciando que o grupo COOH é extremamente
ácido e facilmente ionizável em pH baixo e o grupo amina tem caráter básico,
sendo ionizado em faixas altas de pH. É possível observar também que nas
regiões de pKa (1 e 2) a
glicina tem ação tamponante, ou seja, resiste a variações pequenas de pH em
aproximadamente 1 unidade para mais ou para menos, evidenciando que a glicina é
um bom tampão em pH 2,34 e 9,60, porém não é um bom tampão em pH fisiológico
(em torno de 7,4).
Todos os aminoácidos dipróticos (a
maioria dos aminoácidos) possuem curvas de titulação e ionização parecidos com
o da glicina, porém aqueles triproticos (aminoácidos com grupos funcionais na
cadeia lateral) tem curvas distintas de ionização dos seus grupos, Figura 7.
Figura 7. Curva de titulação dos
aminoácidos triproticos Glutamato (a) e Histidina (b).

Fonte: NELSON; COX,
2014.
É possível
observar que as ionizações dos grupos funcionais dos aminoácidos embora tenham pKas
semelhantes, eles não são idênticos, e esta diferença está relacionada ao
ambiente químico imposto pelos grupos funcionais das cadeias laterais. O grupo
R do Glutamato é facilmente ionizável em pH em torno de 4,25, enquanto a Histidina
tem seu grupo R ionizado em pH 6,0. Observe também que a natureza química do
grupo R, reflete a carga líquida destes aminoácidos.
O aminoácido Histidina oferece uma
ação tamponante em pH 6,0 (pKr),
por este motivo é encontrada em líquidos intra e extracelulares em animais e
bactérias, pois se aproxima do pH neutro (7,0).
Referencias
NELSON, D. L.; COX,
M. M. Princípios de bioquímica de
Lehninger. 6. ed. São Paulo: Artmed, 2014.
DEVLIN, T. M. Manual
de bioquímica com correlações clínicas. 7. ed. São Paulo: Blucher, 2011.
TOY, E. C.; SEIFERT JR., W. E.; STROBEL, H. W.; HARMS, K. P.
Casos clínicos em bioquímica. 3. ed.
Porto Alegre: AMGH, 2016. 480 p.
Não entendi a seguinte parte: "O sistema D e L diz somente respeito à configuração absoluta e não à atividade óptica da molécula". O sistema D e L não estão relacionados com o desvio da luz polarizada e portanto uma atividade óptica? Ou seria apenas uma convenção de nomenclatura?
ResponderExcluir